カイアシ類の体制 ノープリウス幼生の成長
ノープリウス幼生(nauplius larva)とは、甲殻類の幼生であり、形態は成体とはかけ離れています。卵内でノープリウス幼生を過ごし、孵化後には成体と同じ形態となる種は多いですが、カイアシ類を含む種は卵から孵化した個体はノープリウス幼生です。
カイアシ類のノープリウス幼生は6期で構成されます。ノープリウス1期を略してN1と表記することがあります。以降は、その略を使っていきます。N6から孵化するとコペポディット幼体(C1)となり、5回脱皮することでC6、成体となります。(参考記事:「カイアシ類の体制 カラヌス目カイアシ類の成長と体節」)しかし、種によって、特に寄生性はステージを省略することがあり、ヒジキムシ(Pennella)はN2(Wilson 1917)、ツノカクレムシ(Colobomatus)はN5(Izawa 1975)、MytilicolaはN2(Gee & Davey 1986)、ウオジラミ(Caligus)はN2(Ogawa 1992)でノープリウス幼生を終えます。寄生性は生殖に重きを置いており、すぐに生殖能をもつために幼生時期を短縮する戦略なのでしょう。これは、プロジェネシス(progenesis)という幼形進化のひとつです。
ノープリウス幼生は発達具合により二分されることがあります。孵化直後は3対の付属肢、第1触角、第2触角、大顎をもち、体節はなく、単純な形態です。これはオルトノープリウス(orthonauplius)と呼ばれます。N3またはN4をさかいにメタノープリウス(metanauplius)となり、小顎、あるいは顎脚と遊泳脚の原基をもち、体節があります。
成長段階ごとで、ノープリウス幼生の形態は変化します。その変化具合から成長段階を同定することができます。各成長段階の特徴的な形態を、Calanusを例にして下記に記します(Pinchuk 1997;Costanzo etal 2002;古賀 1984)(Fig. 1)。
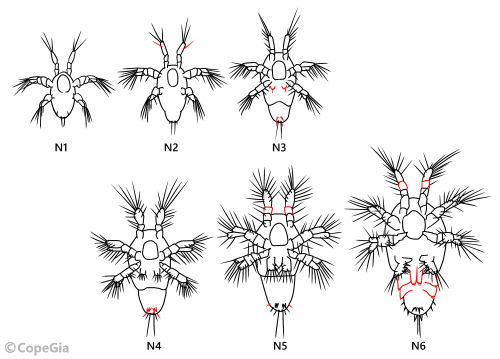
- N1:上唇がよく目立ち、胴体の大半を占める。第1触角の末端節(第3節)に3本の刺毛。途中の節に刺毛はない。大顎の基節、底節に刺毛はない。第2触角内肢末端に3本の刺毛。
- N2:第1触角の末端に4本の刺毛。途中の節に刺毛。大顎の基節に刺毛。
- N3:胴体が2節に分かれる。第1小顎の原基が現れ、1本の刺毛。後部に1本1対の剛毛。大顎の基節に刺毛。第2触角外肢に4本の刺毛。
- N4:後部に2本1対の剛毛。後部側端に2~3本の剛毛。大顎に顎器が現れる。
- N5:第2触角内肢に5本の刺毛。第1触角は4節。
- N6:第1触角が5節。顎脚、第1胸脚、第2胸脚の原基が現れる。
成長段階ごとの形態は、ある程度の種で共通しますが、Calanusと同じカラヌス目(Calanoida)内でも多少の変化はあり、早い時期で遊泳脚の原基をもったり、左右非対称の形態をもったりします(Onbe etal 1988)。他にキクロプス目(Cyclopoida)のOithonaはN2で第1触角末端に4本の刺毛が見られない、N3またはN4で後端刺毛が2対になる(Takahashi & Uchiyama 2007)といった、科あるいは、属、種ごとでノープリウス幼生の成長は異なります。
カイアシ類のノープリウス幼生は種の同定は難易度が高いです。科までしか同定できない場合があります。種の同定には、付属肢の形状や、体形、尾部の後端や側端にある棘や刺毛などの尾部保護装置(caudal armature)を使います。ノープリウス幼生の同定では、古賀(1984)が優れています。Fig. 2~4にそれぞれCalanus sinicus(Calanoida)、Oithona davisae(Cyclopoida)、Epactophanes richardi(Harpacticoida)のノープリウス幼生の写真を載せます。多少の形態の違いが見られます。



文 献
Costanzo G, Zagami G, Crescenti N, Granata A (2002) Naupliar development of Stephos longipes (Copepoda: Calanoida) from the annual sea ice of Terra Nova Bay, Antarctica. J. Crust. Biol. 22 (4): 855-860.
Gee J M and Davey J T (1986) Stages in the life history of Mytilicola intestinalis Steuer, a copepod parasite of Mytilus edulis (L.), and the effect of temperature on their rates of development. J. Cons. Int. Explor. Mer. 42: 254-264.
Izawa K (1975) On the development of parasitic copepoda II. Colobomatus pupa Izawa (Cyclopoida: Philichthyidae). Publ. Seto Mar. Biol. Lab. 22: 147-155.
古賀文洋 (1984) 橈脚類ノープリウス幼生の形態、成体、分類ならびに分化に関する研究. 南西海区水産研究所研究報告 16: 95-229.
Ogawa K (1992) Caligus longipedis infection of cultured Srtiped Jack, Pseudocaranx dentex (Teleostei: Carangidae) in Japan. Gyobyo Kenkyu 27 (4): 197-205.
Onbe T, Hotta T, Ohtsuka S (1988) The development stages of the marine calanoid copepod Labidocera rotunda Mori. J. Fac. Appl. Biol. Sci. 27: 79-91.
Pinchuk A I (1997) Naupliar development of Metridia pacifica (Copepoda: Calanoida) reared in the laboratory. J. Crust. Biol. 17 (2): 343-351.
Takahashi T, Uchiyama I (2007) Morphology of the naupliar stages of some Oithona species (Copepoda: Cyclopoida) occurring in Toyama Bay, southern Japan Sea. Plankton. Benthos Res. 2 (1): 12-27.
Wilson CB (1917) North American parasitic copepods belonging to the family Lernaeidae with a revision of the entire family. Proc. U.S. Natl. Mus. 53: 1-150.