カイアシ類の歯
カイアシ類は数mmサイズの甲殻類で、海や川、湖、池、ごく一部は土壌中にも生息している。多くは浮遊生活をするプランクトンだが、底生性(ベントス)、寄生性も存在する。
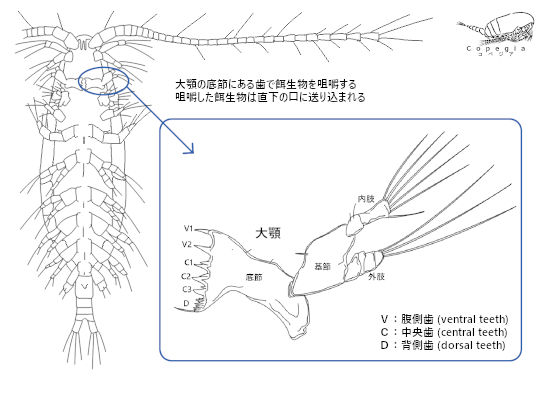
そんなカイアシ類だが、餌生物を摂食する際に、他の動物のように歯を用いて咀嚼をする。咀嚼歯は大顎にある。カイアシ類を腹側から見た際に、口の近くに位置する。節足動物全般に、大顎や小顎といった付属肢は、遊泳脚のような形態から変形したもので、「顎」と言えど、口内の器官というわけではない。カイアシ類の顎は遊泳としても機能するため、世間一般的な「顎」のイメージと、かけ離れているが、他の動物、たとえばオオスズメバチがもつような強靭な大顎と相同器官である。
大顎は、底節、基節、内肢、外肢からなり、咀嚼歯は底節にある。咀嚼歯は複数あり、腹側歯(V歯)、中間歯(C歯)、背側歯(D歯)がある(Fig. 1)。多くの場合、V歯は頑丈で鋭利になっており、C歯はV歯より尖ってなく、D歯は微細な歯が並んでいる。さらにその外側に剛毛(out dorsal setae)がある。
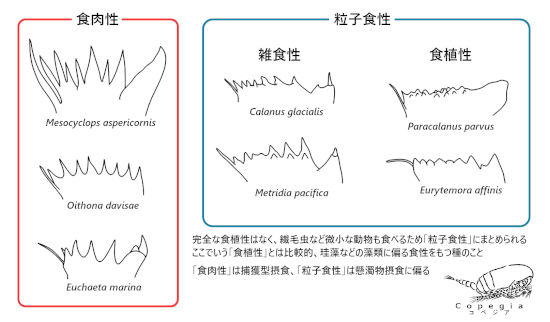
肉食性の動物には鋭い牙があり、草食性には大きな臼歯があることは有名なことだろう。肉を噛みちぎるために犬歯が発達し、草をすりつぶすために臼歯が発達する。実は、カイアシ類でも、そういった形態の特殊化が見られる。例として、7種の咀嚼歯をFig. 2に示した。甲殻類幼生や鞭毛虫を食べるようなOithona属とEuchaeta属の咀嚼歯はV歯、C歯ともに鋭利になっている。昆虫のボウフラを食べるMesocyclops属にいたってはどの歯も鋭く尖っている。珪藻やその他の植物プランクトンを食べるCalanus属やParacalanus属は鋭利な歯は目立っておらず、背の低い歯が並んでいる。しかし、純粋な食植性は少なく、微小な鞭毛虫や繊毛虫を食べることがある。食植性というよりかは、運動しない微小な生き物、すなわち粒子食性と呼ぶのが好ましい。その中で、比較的に食植性という位置づけがある。
厚い被殻(ひかく)をもつ珪藻は、カイアシ類にとっては硬すぎて歯が破損してしまうかもしれない。そこで「オパール歯」と呼ばれる、歯にシリカ質を沈着させ、強固化した種はいくつか知られている(Michels et. al. 2012)。中には、さらに特殊化し、オパール歯となったV歯が牙のような形状となり、さらにV歯の先から毒を出すHeterorhabdus spinifronsという種がいる(Nishida and Ohtsuka 1996)。まさにヘビの毒牙である。毒の成分は未だに不明だが、大顎の付け根付近にある上唇から分泌され、筒状になったV歯を伝わって、餌生物へ毒を注入するようだ。進化とは、ここまで特殊化させてしまうのだから驚きを隠せない。
食性によって咀嚼歯の形状が適応していくことだが、咀嚼歯の形状から食性を推定することは可能である。Itoh(1970)は歯の大きさや間隔の広さなどのパラメーターを使って、指数化する式を導いた。それによると、指数が500以下ならば食植性、900以上ならば食肉性と推定できるとのことだ。ここでは触れる程度にするが、興味ある方は、文献を見ていただきたい。
文 献
Dalpadado P, Yamaguchi A, Ellertsen B, Johannessen S (2008) Trophic interactions of macro-zooplankton (krill and amphipods) in the Marginal Ice Zone of the Barents Sea. Deep-Sea Research II 55: 2266-2274.
Ferrari F D, Orsi J (1987) Oithona davisae, new species, and Limnoithona sinensis (Burckhardt, 2012) (Copepoda: Oithonidae) from the Sacramento-San Joaquin Estuary, California. J. Crustacean Biol. 4 (1): 106-126.
Fuentes-Reinés J M, Suárez-Morales E, Granados-Marínes C E (2017) First occurrence of Mesocyclops aspericornis (Daday, 1906) (Copepoda: Cyclopoida) in northern Colombia. Check List 13 (2): 2076.
Giesecke R, Gonzáles H E (2004) Mandible characteristics and allometric relations in copepods: a reliable method to estimate pray size and composition from mandible occurrence in predator guts. Revista Chilena de Historia Natural. 77: 607-616.
Gurney R (1931) British fresh-water copepoda. Vol. 1. The Ray Society, London. 238 pp.
Itoh K (1970) A consideration on feeding habits of planktonic copepods in relation to the structure of their oral parts. Bull. Plankton Soc. Japan. 17: 1-10.
Michels J, Vogt J, Gorb S N (2012) Tools for crushing diatoms – opal teeth in copepods feature a rubber–like bearing composed of resilin. Sci. Rep. 2: 465.
Nishida S, Ohtsuka S (1996) Specialized feeding mechanism in the pelagic copepod genus Heterorhabdus (Calanoida: Heterothabdidae), with special reference to the mandibular tooth and labral glands. Mar. Biol. 126: 619-632.
大塚 攻, 西田周平 (1997) 海産浮遊性カイアシ類(甲殻類)の食性再考. 海の研究. 6 (5): 299-320.
Ohtsuka S, Shimozu M, Tanimura A, Fukuchi M, Hattori H, Sasaki H, Matsuda O (1996) Relationship between mouthpart structures and in situ feeding habits of five neritic calanoid copepods in the Chukchi and northern Bering Sea in October 1988. Proc. NIPR Symp. Polar Biol. 9: 153-168.
Sullivan B K, Miller C B, Peterson W T, Soeldner A H (1975) A scanning electron microscope study of the mandibular morphology of boreal copepods. Mar. Biol. 30 (2): 175-182.